














(In this post I'm only addressing the specific points made in this Q&A. Magenta numbers identify points I have concerns about. As always, I'm not asking readers to take my word for anything; if you have doubts you should look up the information and check my calculations for yourself.)
Question:
Some people have questioned whether the DNA was sufficiently cleaned by your technique using gel electrophoresis, to separate it from other molecules. Do you feel this is a valid concern?
Answer:
Our DNA extraction and purification protocol begins with washed cells, pelleted from media. These are then subjected to a standard DNA extraction protocol (1), which included multiple phenol chloroform steps to remove impurities, including any unincorporated arsenate (As) (2). After this, the DNA was electrophoresed, further separating the DNA from impurities (3, 4). Any residual As from the media would have been removed by washing the cells prior to extraction (5) and by partitioning into the aqueous phase (6) during the 3 phenol:chloroform steps in the extraction. If As was incorporated into a lipid or protein it would have partitioned into the phenol, phenol:chloroform, or chloroform fractions (7). Additionally, DNA extracted in this manner on other samples was also successfully used in further analyses, including PCR (8), that require highly purified DNA (9).
The arsenic measured by NanoSIMS in the gel band is consistent with our other measurements and another line of evidence.
Our radiolabeled 73AsO43- experiment showed that of the total radiolabel associated with the cell pellet 11.0 % ± 0.1 % was associated with the DNA/RNA fraction (10). This indicated that we should expect some arsenate of the total pool associated with the nucleic acids (11). To interpret these data, we coupled our interpretation with our EXAFS evidence suggesting that intracellular arsenic was As(V) bound to C (12), and was not free in solution as an ion. This suggests the As is in, an organic molecule with bond distances consistent with a chemical environment analogous to phosphate (Figs. 3A, S3 "bond lengths" table). Further supporting our interpretation of the previous mentioned two analyses, we used a third line of evidence from NanoSIMS, a completely different technique from the other two. We find elemental arsenic (as measured by NanoSIMS) associated with the gel band that is more than two times the background in the gel (13). Based on the above discussion, we do not feel this is a valid concern.
My concerns:
(1) The DNA extraction procedure included only some of the components of a standard DNA extraction protocol. First, only a single ethanol precipitation was done, whereas getting relatively pure DNA requires at least two rounds of extraction and precipitation. Second, the pellets were not washed, so that unincorporated arsenate (or phosphate) present in the aqueous fractions may have been precipitated with the DNA, and may also have been present in the alcohol supernatants contaminating the pellets. Third, no column clean-up step was done.
(2) This statement implies that unincorporated arsenate in the cell lysate would partition into the phenol and chloroform. This seems a priori improbable, as arsenate is very soluble in water. No controls were done to find out how unincorporated arsenate or phosphate would partition in these extractions.
(3) Gel electrophoresis can remove impurities but it is not guaranteed to do so. Any impurities that migrate at a similar rate to the DNA, or are electrostatically associated with it, will be present in the gel slice. Any impurities that diffuse into the gel buffer may become distributed throughout the gel. Any impurities already present present in the agarose or gel buffer will also be in the gel slices. These concerns are strengthened by the failure to purify the DNA away from the gel slice (see (13) below).
(4) No control was performed for non-covalent association of arsenate (or phosphate) with DNA. In another control extraction, arsenate and lysis solution should have been mixed with previously purified DNA from E. coli or other phosphate-grown cells, to see if any arsenic co-purified with the DNA.
(5) No control was performed for the effectiveness of this washing. E. coli or other cells grown in the absence of arsenate should have been mixed with the +As/-P medium and then subject to the same washing and extraction steps.
(6) Here arsenate is predicted to partition into the aqueous phase. Is this the same arsenate that partitioned into the phenol and chloroform in (2) above?
(7) In the ICP-MS analysis presented in Table S1, almost all the arsenic did partition into the phenol phase, and almost as much arsenic was present in the phenol fraction of the phosphate-grown cell (4725 vs 3683 ppb). In fact, the aqueous phase of arsenate-grown cells contained no detectable arsenic at all, even though this is the fraction from which the arsenic-containing DNA was precipitated.
(8) The online Methods say that DNA from all growth conditions, worked fine in the PCR reactions used for the phylogenetic analysis. This strongly suggests that the DNA from arsenate-grown cells has a normal phosphorus backbone. The polymerases used for PCR have very high fidelity and would not tolerate substitution of arsenic for phosphorus.
(9) PCR does not require purified DNA; it even works very well on whole-cell lysates.
(10) See point (7)
(11) But this pool is expected to contain all of the water-soluble constituents of the cell. The elemental analysis report that the arsenic was bonded to carbon doesn't mean that it is bonded to DNA. And most of this arsenic partitioned into the phenol phase - is it thought to be lipid? If so, maybe the arsenic is bonded to C in lipids.
(12) Wait! IANAC (I am not a chemist), but if the 'intracellular arsenic was As(V) bound to C' then it couldn't be arsenic incorporated into DNA or RNA, as it would then have to be bound to O in DNA's diester backbone.
(13) Let's think more about the arsenic in the gel bands. The whole gel slices were assayed (the DNA was not purified away from the agarose); since the gel is 1% agarose and a gel slice is unlikely to weigh less than 100 mg, each slice would contain at least 1 mg of agarose. The DNA bands in Fig. 2A are unlikely to contain more than 1 µg of DNA, probably less for arsenate-grown DNA in lane 2). Thus we can generously assume that 99.9% of the carbon in each DNA sample came from the agarose, and no more than 0.1% from the DNA. According to the figure legend and the numbers at the bottoms of the gel lanes the arsenate-grown sample had 13.4 atoms of arsenic per 10^6 atoms of carbon. This is 13.4 arsenic atoms per 1000 DNA carbons. Since A, G and T nucleotides contains 10 carbons and C has 9, this is 13.4 arsenic atoms per 102.6 nucleotides, or about 26 per 100 base pairs. That's quite a lot of arsenic. Even more surprising, the phosphate-grown sample had 6.9 arsenics per 10^6 carbons, which would be about 14 arsenics per 100 bp. The gel blank had even more arsenic, and three times as much phosphorus. This strongly suggests that the gel was contaminated with both arsenic and phosphorus, perhaps introduced with the DNA samples. Until such contamination can be ruled out, the two-fold higher arsenic concentration and three-fold lower phosphorus concentration associated with the arsenic-grown DNA sample cannot be seen as significant.
Question:
Others have argued that arsenate-linked DNA should have quickly fallen apart when exposed to water. Could you address this?
Answer:
We are not aware of any studies that address arsenate bound in long chain polyesters or nucleotide di- or tri-esters of arsenate, which would be directly relevant to our study. Published studies have shown that simple arsenic esters have much higher hydrolysis rates than phosphate esters (1-3). The experiments published to date have specifically looked at the exchange or hydrolysis of alkyl tri-esters of arsenate [Eqn. 1] and alkyl di-esters of arsenite [Eqn. 2]:
OAs(OR)3 + H2O → OAs(OH)(OR)2+ ROH [1]
OAs(OH)(OR)2 + H2O → OAs(OH)2(OR) + ROH [2]
where R = methyl, ethyl, n-pentyl and isopropyl. Reference 2 demonstrated that the hydrolysis rates for these simple alkyl triesters of arsenate decreased with increasing carbon chain length (complexity) of the alkyl substituent (methyl > ethyl > n-pentyl > isopropyl) (14). No work has been done on the hydrolysis rates of arsenate-linked nucleotides or other biologically relevant moieties.
If the hydrolytic rate trend reported in Ref. 2 continues to larger-weight organics, such as those found in biomolecules, it is conceivable that arsenate-linked biopolymers might be more resistant to hydrolysis than previously thought (15). The small model compounds investigated in Refs. 1-3 are relatively flexible and can easily adopt the ideal geometry for water to attack the arseno-ester bond (16). Arsenate esters of large, bio-molecules, however, are likely to be more sterically hindered leading to slower rates of hydrolysis (17).
This type of steric constraint on reaction rate accounts for the wide range of rates seen in the behavior of some phosphate linked nucleotides. In small ribozymes, the phophodiester linkages at the site of catalysis can be hydrolyzed on the order of tens of seconds (with a chemical rate of 1 s-1). This rate enhancement is achieved by orienting the linkage for in-line attack by a nucleophile (an adjacent 2' hydroxyl group). Moreover, the autodegradation patterns are consistent with specific base composition. On the other hand, the hydrolysis rates for phosphodiester bonds in A form duplexes of RNA are many orders of magnitude slower, because these linkages cannot access easily the geometry necessary for hydrolysis.
The rates in DNA may be much slower than model compounds because of the geometrical constraints imposed upon the backbone by the helix (18).
The kinetics of the hydrolysis of arsenate-linked biopolymers is clearly an area where more research is warranted.
My concerns:
(14) Again IANAC. But note that these are hydrocarbons and thus quite hydrophobic, especially the pentyl chain (5 carbons) and the isopropyl chain (3 branching carbons). Attaching three of these by ester bonds to the arsenate effectively surrounds the bonds with hydrophobic shells that exclude water. Thus it's not surprising that the hydrolysis reaction occurs less often.
(15) But if the increased stability described in Reference 2 (yes, I looked at this paper, and I'm trying to get hold of the 1870 reference too) is due to increasing hydrophobicity of the ester bonds' environment, then the effect will not extrapolate to long hydrophilic biological molecules such as DNA.
(16) Not if they're surrounded by a hydrophobic shell, and probably also associating with each other to reduce the shell's exposure to the aqueous solvent.
(17) DNA has its backbone on the outside of the double helix, and the entire molecule is quite hydrophilic.
(18) Assuming the overall structure of arsenic bonds in DNA is like that of phosphorus bonds in DNA, shouldn't any stability-enhancing geometrical constraints of the DNA structure be experienced by both phosphorus and arsenic bonds? We would then still expect arsenic bonds in DNA to be 100-fold less stable than phosphate bonds.
Question:
Is it possible that salts in your growth media could have provided enough trace phosphorus to sustain the bacteria?
Answer:
The data and sample labeling in Table S1 has caused some confusion. To clarify, for every experiment, a single batch of artificial Mono Lake water was made with the following formulation: AML60 salts, no P, no As, no glucose, no vitamins. Table S1 shows examples of ICPMS measurements of elemental phosphorus (~3 µM) and arsenate made on this formulation prior to any further additions (19, 20). Then we added glucose and vitamins for all three treatments and either As for the +As treatments or P for the +P treatments. The P measurements made on the medium after the addition of sucrose and vitamins and after addition of As were also ~3 µM in this batch. Therefore, it was clear that any P impurity that was measured (~3 µM, this was the high range) came in with the major salts, and that all experiments contain identical P background (including any P brought in with the culture inocula).
In the Science paper, we show data from one experiment of many replicated experiments that demonstrates no growth of cells in media without added arsenate or phosphate (Figure 1). These data clearly demonstrate that strain GFAJ-1 was unable to utilize the 3µM P to support further growth in the absence of arsenate (21). Moreover, the intracellular P content determined for the +As/-P grown cells was not enough to support the full requirement of P for cellular function (22).
Note on culturing: All experiments were initiated with inocula from sustained +As/-P conditions. Prior to the experiments, the cells had been grown long term, for multiple generations from a single colony grown on solid media with no added phosphate. Before this, they were grown as an enrichment for more than 10 transfers and always into new medium that was +As/-P. We therefore feel that there is not significant carry-over of P. We also argue that there would not have been enough cellular P to support additional growth based on an internal recycling pool of P (23).
My concerns:
(19) The two batches of AML60 salts assayed contained 3.7 and <0.3 µM P, and the single batch of cell wash solution contained 7.4 µM P. Given this variability, the similarity of the two batches of -P/+As medium doesn't inspire much confidence.
(20) Was no effort made to identify and eliminate the source(s) of this contamination?
(21) Agreed, with the proviso that the media be tested and shown to be identical except for the phosphate and arsenate. But this wouldn't mean that arsenic replaced phosphorus in any biological molecules in GFAJ cells, just that the cells needed arsenate for something.
(22) Did this calculation take into account the very high carbon content of the poly-hydroxybutyrate granules in these cells? PHB can account for up to 90% of the dry weight of phosphate-starved cells, and its carbon will skew estimates of C:P ratios.
(23) This assertion is not supported here by any evidence, and it is contradicted by the Makino et al. (2003) reference cited by the Science paper (ref. 13). These authors found that 10% of the P in phosphate-limited E. coli is in the DNA and the rest is in RNA and other cellular components. Using this value and your own estimate of genome size (3.8 Mbp), ~3 µM P is sufficient to account for the observed growth in the -P/+As medium. Here's the calculation:
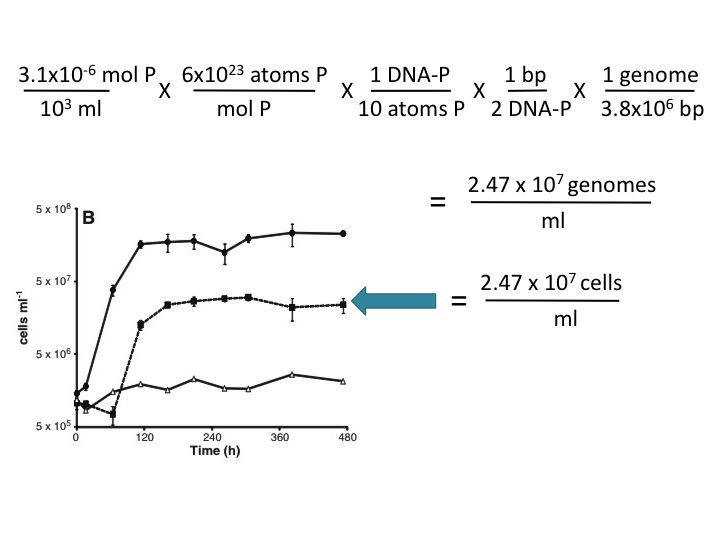
I don't *quite* get the 1 DNA-P/10 atoms P step.
ReplyDelete1 DNA-P/10 atoms P
ReplyDeleteis a rough assumption that 10% of the P in the cell is used for the DNA, the 90% left is used for other molecules.
I am a scientist (not a biologist) and have followed this whole debate with interest. There is one question you do not seem to have addressed (correct me if I'm wrong): if this is such a flawed study, as you claim it is, how do you explain its acceptance through the peer review process? Are you suggesting a corrupt process? Favoritism? Editorial bias? Lack of scientific rigor? Poor selection of peers for the review? Your rebuttals have received so much attention, and perhaps deservedly so, but I have to question the widespread insinuations of wrongdoing, if only because they are so vehement. I understand that this study makes claims that if substantiated would fundamentally change our understanding of life, which gives it unusual weight. I also understand that there are methodological factors at issue which can only be fairly evaluated by peers. So how do you explain the decision by your peers to accept its methodology and allow this paper to be published in a highly respected journal? Is it possible that the biases are represented by the dissenters? Or that perhaps there are honest differences among experts regarding preferred methodologies that might explain their acceptance of it while others disagree? No offense intended, but I wonder how a paper as flawed as you claim it is could pass through the review process.
ReplyDeleteIs the 10% to DNA a commonly accepted number? Early Dr. R mentioned half to the nucleus, half to RNA and other molecules.
ReplyDeleteWhen I read the FAQs yesterday night I could not sleep. Finally I had a reliable assay for the purity of DNA! Who would have thought it would be as simple as a PCR?
ReplyDeleteSo instead of going to bed, I went out to test a theory of mine and performed PCR on a variety of samples:
a leaf
some swamp water
a hazelnut
a feather
some animal 'leavings'
a piece of moss
As is suspected, they all came back positive; they were all composed solely of purest DNA! Remember: PCR works = no protein, no lipids, no salts, polysaccharides...!
I've already drafted a paper entitled 'Southern England: A unique ecosystem composed entirely of DNA'. I expect it to be published shortly. I'm very exited!
Justin Cayce
How can flawed papers pass peer-review? Easy: overworked reviewers, heard about it at a conference and didn't read it properly, the parts in your field of expertise were ok, etc. etc. Flawed methodology passing peer-review happens all the time, you can't prevent it. But you can correct it, that's the main point. Pre-publication peer-review reduces the flaws, post-publication review can eliminate them. Both is required and helps.
ReplyDeleteand don't forget that the most negative reviews can be overruled by eager editors. true for most big impact factor journals.
ReplyDeleteRosie: well done, as usual. keep on fighting for the right of science to be performed in a proper way. it deserves it.
@scientist(not a biologist)
ReplyDeleteBy saying that you '...understand that this study makes claims that if substantiated would fundamentally change our understanding of life...', I think you have touched on one of the main issues of the whole arsenate saga. Aside from the glaring omission of necessary controls, poorly presented and at times contradictory data (e.g. %As and P vs. As/C and P/C ratios), what irks many biologists and biochemists that I have spoken to is the fact that the conclusions were massively oversold to the public.
Arsenate DNA would not revolutionise biology. If it existed, it would be interesting to see how it is stabilsed and synthesised, but otherwise there is not much in it. The existence of a bacterium that could incorporate As into DNA would be interesting, but As has been known to be able to replace P in biochemical reactions for a very long time indeed. Also, this bacterium is firmly rooted in the tree of life as we know it, and has nothing to do with a 'shadow biosphere' (i.e. a domain of life that is fundamentally different from life as we know it). This is utterly 'life as we know it'.
Many biologists feel that NASA has hyped up the (tenuous!) conclusions from the paper to generate a media storm. Of course, NASA may have acted in good faith and actually believed that theirs was a major discovery.
It all boils down to the question 'How far can scientist go to spice up their findings without being guilty of misleading non-experts?'.
As scientist we are all very aware of the need to 'sell' our science, but a lot of people believe that a line was crossed in this case. The fact that the study was (in the eyes of many) poorly controlled did not help.
Scientist(biochemist)
I am not a biologist but a scientist who is quite interested in this #arseniclife debate.
ReplyDeleteI do appreciate that you are taking your time to clarify your view and answer to this Q&A. However I wonder why if you are so skeptical, you don't simply try to redo their experiment with your own group and your own techniques. It will be more convincing to show by the experiment that they are wrong than to write thousands of lines on a blog.
@F. Marchis:
ReplyDeleteusually this could be the case. However, here we are faced with unproved science published in a big journal with way too much hype.
The authors are responsible not only for data they publish (which, per se, could be accepted in a much smaller journal as preliminary findings) but also for the flawed and misleading interpretation they give.
Every paper has some pros and cons, some things could have been done better or a missing control, but the story about GFAJ-1 (which stands for Give Felisa A Job, as a collaborator of the main author said, let us remember this) has NO controls. The authors argument about the controls in the same way as we argument about what can be wrong in their study. With the difference that this IS their study, and it is their responsability, once again, to prove it before publish it.
@F. Marchis: Redoing the experiments properly would be a lot of work. Not only is the analytical chemistry far outside of my expertise, but I have no funding for this work. Take a look at Jim Hu's post about why he won't be redoing these experiments: http://dimer.tamu.edu/simplog/archive.php?blogid=3&pid=6947
ReplyDeletePointing out flaws in other people's work is not lazy science - it's a vital part of what makes the process work. We always need to carefully examine the possible flaws BEFORE we go on to the next step.
Unfortunately I can't see that the information Wolfe-Simon et al provided answers the raised question as discussed above, in some cases they are aware of that.
ReplyDeleteAnd what does this mean: "For all of us, our entire team, what this was like was unimaginable."
If it refers to the discovery, it is _exactly_ what they imagined and have discussed before. If it refers to the reactions, it is exactly what they should have expected/wanted/accepts.
As for this post, how the As goes out in the washes are contradictory as described; however I feel that this may be the point.
But this may be a simple type error from the response authors: "IANAC (I am not a chemist), but if the 'intracellular arsenic was As(V) bound to C' then it couldn't be arsenic incorporated into DNA or RNA, as it would then have to be bound to O in DNA's diester backbone." The table 3 in the paper gives the first and main EXAFS peak in fig 3 as roughly As-O with order 4, which would correspond to phospate as in the DNA backbone, I believe. But IANAC either.
Btw, this WP article has elevated a comment on the first criticism of this blog, which unfortunately is from what would in general web threads be considered "a concern troll". I.e. complaining about tone diluting the message.
The big question is where is she heading to now... The periodic table is huge...
ReplyDeleteAntimony is an obvious choice (moving downwards).
C substituted for Si is another direction...
H is troublesome. There I would be undecided, really.
O - S - Se is seemingly not promising, but hey! there can be a permutation here.
Yep. They should have run controls with 'As-free' grown bacterial samples doped with As to see how well their purification methods worked.
ReplyDeleteCan everyone please get over the fact that NASA and Wolfe-Simon overhyped these findings with the press conference? So what? That has no bearing on the actual science. Also, a lot of people have criticized the use of the term "astrobiology" in promoting the press conference, but this is a legitimate and often used term for these types of studies, there is even a journal called "Astrobiology" that often publishes research on extremophilic bacteria, so noone was trying to be misleading with this term. No life has been found on other planets yet but the field of astrobiology studies unexpected forms that life *could* take. Personally I don't like the term but it has been in use for a long time before NASA's press conference.
ReplyDeleteI completely agree with Dave. Who cares if some scientists completely distort the importance of their supposed find to get some attention and funding? What's the big deal? It's not like the money could be better spent on research that is supported by actual data or anything. Get a life, people! If anything, science will advance quicker if we don't have to bother with proper experiments anymore.
ReplyDeleteMy point was that people inferred it was going to be an anouncement about finding life on Titan, and that contributed a lot to the overblown press reaction. I don't think NASA anticipated that.
ReplyDeleteDave: Overhype is one thing, overhype of bad science is quite another. If NASA and Wolfe-Simon had quietly published this paper it clearly would not have triggered such as blistering response. However, since NASA told us we should be taking an eraser to our biology textbooks..... they have to expect stringent criticism if their scientific case is lacking.
ReplyDeletehttp://astrobiology.nasa.gov/articles/thriving-on-arsenic/
Dr Redfield: excellent analysis! (as was your previous one on the subject). What those commenters with the "get a life" attitude don't realize is that a researcher(s) her/himself(themselves) who submit a paper for publication should be the one(s) doing such a stringent analysis of their work. Not doing so is, under any circumstance, a blatant lack of intellectual honesty. A scientist always must be his/her own greatest critic.
ReplyDeleteThe situation is aggravated further because publishing in Science isn't supposed to be an easy task because of the large editorial and reviewer demands on subject novelty and good experimental design. That the second demand wasn't met is obvious, as well as that somebody must have pulled some strings to get the paper to overcome that obstacle. That by itself is scandalous, and the matter is made worse by the instigation of a media hype over what is turning out to be no more than a gaffe on NASA's and Wolfe-Simon's team's side. However one looks at it, that's terrible publicity for science.
Their conclusions are a valid interpretation of the data that they saw from multiple different lines of evidence. What research paper is ever 100% certain in its conclusions? You have a lot of "improbability arguments" against their conclusions but little else. The personal attacks like "bad scientists", "intellectual dishonesty" and "pulling strings" are baseless, borderline libelous, and do not contribute anything positive to the discussion.
ReplyDeleteBy the way, I agree that the NASA press conference was really ill-advised considering that this study represents the very early stages of research into this topic. Like I said, interpretation is never 100% especially for something this new. They really shot themselves in the foot by doing that (for now).
If you want to publish in Science, yes, you (usually) have to have more certainty than what Wolfe-Simon et al showed. That's why I find their publication highly suspicious (as many other scientists do). As Dr Redfield said in her previous critique, what they present would be enough to send a graduate student back to the lab to do more work, this isn't what you expect from somebody with a NASA grant.
ReplyDeleteYour "borderline libelous" argument is an "improbability argument". Have you ever participated in a journal club?
Ribosome, I'm not sure I follow the logic of the second paragraph of your last post, but does participation in journal clubs usually involve posting unfounded accusations of "pulling strings to get published" on the internet?
ReplyDeleteEveryone needs to calm down, only further research will really answer this question and it surely will happen sooner rather than later.
Awesome post! just like your previous one. This study was shabby science, and exposing shabby science for what it is is a valuable public service -- especially where a study as overhyped as this one is concerned.
ReplyDelete@anonymous: Again, I'm not against debating the science, especially when more data is produced to either support or deny the claim. If the authors conclusions are wrong it doesn't change the fact that they were a legitimate interpretation given the data that they had from multiple lines of evidence. It doesn't mean that there was any inpropriety in how the paper got published. Do you deny that only more research will truly answer the question? I'm done arguing here, everyone has already made up their minds much like you accuse Wolfe-Simon et al of doing. I would be very curious to know what you all will have to say if their findings are substantiated by more research. I will leave you all to your echo chamber now.
ReplyDeleteI am an optical physicist and have seen the whole race for high impact papers at close quarters. dr. redfield's arguments may prove true as far as this particular study is concerned. but she has crossed the line in peer criticism at several levels. dr. wolfe is a motivated if not naive young scientist.
ReplyDeletedr. redfield, if her intentiions were to provide criticism has clearly lacked the judgment that a senior scientist like her should demostrate. irrespective of what this survey shows, we come across crtics like her and it only makes academia
a thanksless place for young scientists.
dr. redfiled might have proven her point but she has done a massive disservice to a young scientist and other young scientists in her community. this is not the first time that an eager new scientist has done a mistake but by choosing an echo chamber like blogsites, what redfield did is deplorable. if you are so good at what you do, you should provide the arguments in person, or in a conference, or in a paper. for a senior scientist she shows very little judgement. i just hope this new area of astrobiology doesnt suffer from thickheads like her.
This part of the OP interested me, re that either P or As had to be added for the bacteria to grow:
ReplyDelete"(21) Agreed, with the proviso that the media be tested and shown to be identical except for the phosphate and arsenate. But this wouldn't mean that arsenic replaced phosphorus in any biological molecules in GFAJ cells, just that the cells needed arsenate for something."
That cells needed arsenate for something would be an interesting finding, I'd guess. And cells need As for something that P does is also implied by this finding.
Is this an area of potential agreement between the original article authors and critics?
(IANAS, BTW, so YMMV)
Brian, if you are not a scientist (IANAS) that was a very astute post. I'm not convinced that the bacteria is replacing P with As in DNA (maybe a small % of it) but there is pretty good evidence in the paper that the bug uses As to fill some biochemical need that is usually met by P.
ReplyDeleteFunny, right after I noted a "concern troll" comment the behavior is repeated.
ReplyDelete"everyone has already made up their minds much like you accuse Wolfe-Simon et al of doing" Are you kidding, it is _precisely_ because we are uncertain of the conclusions that we critique the paper.
Now you can't promote your theory (here standard CHNOPS) by critiquing or, better, test for rejecting another. But you can analyze a putative falsification of yours, which is the situation here. So far the evidence isn't compelling for rejecting CHNOPS.
Well, I have already commented on the category. But this:
ReplyDelete"choosing an echo chamber like blogsites"
is not relevant, not even correct. The "echo chamber" is evaluating this as good, nay better, peer review than "magazine only". Google it.
[I can sympathize with the idea that harsh criticism can be counterproductive for a community. But I would demand statistics before I consider it seriously. Anecdote isn't data.]
Does anyone understand what Torborjn is talking about?
ReplyDeleteOne way or another this will be confirmed or refuted by further work, and all of the opinions on the internet will be irrelevant. Its published in Science, that doesn't make it true, but its enough for me to wait and see what the follow up work reveals. There is some compelling evidence in the paper for many of its claims.
@Torbjörn Larsson, i review for these journals and magazines in question. i stand by what I said. No respectable scientist has taken this route of lashing on a young scientist in public. thats how the decorum of science works. Dr. Felisa might have done a mistake, but what this blogs author is doing is public assault. she has drooped to a level worse than just doing a scientific error. if blogs and public commenting is the reason for that, i shoud just say your community may not be ready for it yet. open sourced publications like arxiv have set a great example of quick dissemination and critical process. Your and this author's choice of words, choice of medium, lack of understanding of what affect an open washing of dirty laundry will have on the community utterly disappoints me. if you talk about funding, which this author mentions, i will love to see if acerbic fighting like this will lead to a single new student or funds in astrobiology. the funding reviewers and peer reviewers like me are watching the terms of this argument. most of my colleagues in private consider this style to be bad and when the time comes it will be made clear.
ReplyDeleteAnonymous: somehow you seem to forget that part of the scientific process is open criticism and analysis of published results. If I were a graduate student or a posdoc, I wouldn't want you supervising my work, as you would be likely to push me to publish incomplete research and obviate stringent controls. Your anonimity and your relaxed attitude towards good experimental methodology make me seriously doubt your supposed work as a reviewer. Your acerbic defense of sloppy publishing, your talk of respectability behind the veil of anonimity and your implied threat of blocking funding approval (how respectable!) make me rather suspect some kind of involvement with Wolfe-Simon's work.
ReplyDelete"...for the phylogenetic analysis. This strongly suggests that the DNA from arsenate-grown cells has a normal phosphorus backbone. The polymerases used for PCR have very high fidelity and would not tolerate substitution of arsenic for phosphorus."
ReplyDeleteRosie, could PCR work if only a small proportion of P was substituted? What's the reason fro saying that PCR polymerases wouldn't tolerate arsenic substitution?
@anonymous (the one at 3:17 pm)
ReplyDeleteI think that the 16S rRNA segment they amplified was about 1500 bp long. If less than about 0.1% of the backbone phosphoruses were replaced with arsenic then the PRC polymerase would probably have had enough arsenic-free template DNA to copy.
The reasons to think that DNA polymerases would not be able to use arsenic-containing templates are spelled out in the post that follows this one.
Here's a question. Isn't it customary/expected that if you publish a letter in the journal Science, and you want to put it on your CV with the Science citation, you should specify that the work in question is in fact a letter and not a full blown article?
ReplyDeleteI mean, if you saw this on someone's CV:
Redfield R. 2009. Looking to bacteria for clues. Science. 325(5943): 946 PDF
you'd be inclined to think its a real paper, right? A letter would be expected to appear more like:
Redfield R. 2009. Looking to bacteria for clues. Letters to Science. 325(5943): 946 PDF
or
Redfield R. 2009. Looking to bacteria for clues. Science (letter). 325(5943): 946 PDF
Am I off base here? Well Rosie at least you provided the link for the PDF so its not intellectually dishonest or anything (heavens no).
By the way, congrats on your upcoming NEW SCIENCE PAPER that you will be able to put on your CV if your latest letter makes it in.
@ Dave: That Letter isn't on my CV, though it is on the list of research I've published, on the 'What we've done' page of our web site. Sorry you were misled.
ReplyDelete@Rosie, OK thanks for clarifying I thought that was a copy of your CV but a website is not really used for academic purposes.
ReplyDelete^
ReplyDeleteDon't be a dick, Dave.
Dave, professors and scientists that are far more reputable than you have personal and lab websites with their CV's posted.
ReplyDeleteThanks Rosie for keeping up the fight, even with jerks like Dave around. People are getting caught up in the delivery and not the message (i.e. the science).
ReplyDeleteSomeone along the way in the comments asked what us skeptics would think if this result is validated? That question exposes a key misunderstanding of the scientific method that is unfortunately shared by the general public. That the authors conclusions are poorly supported by their data is a fact that won't be changed by subsequent work.
Another person asked how such a flawed paper could have gotten past peer-review. Peer review is imperfect for many reasons but is the best system that we know of for screening shoddy science. Bad papers do get through sometimes so the publication of a paper is not a guarantee that it is good science (conclusions following from data).
See the criticism of Wolfe-Simon paper on Facult of 1000:
ReplyDeletehttp://f1000.com/6854956
@Anonymous at 10:45, my last post was meant to be sarcastic so yes I realize quite well that scientists often post their CV online. I also realize that others see these CVs and take them at face value because its impossible to check every entry to make sure the person isn’t being *deliberately misleading*. Thank you for validating my premise.
ReplyDeleteAs for my point in bringing it up, I think it goes to motive. Let’s leave it at that.
@Anonymous at 12:33, it would be a lot easier to avoid getting lost in the delivery if the delivery wasn’t so over-the-top personal, vehement, and insulting. I’m especially referring to the nature of the quotes that were given to anyone in the media who was willing to put a “microphone” in front of Dr. Redfield. Yes, it does dilute your message if you do business this way. It was true before the internet and it’s still true now, in all places except the cesspool that is the blogosphere.
@Dave,
ReplyDeleteThere are a lot of reasons why some screaming is warranted in this situation and I'm glad there are people like Rosie that are willing to do so. Take a look at the news agencies that are still publishing stories that are clearly spoon-fed from the NASA press machine. But thanks to blogs like Rosie's, most news is taking a more balanced view of this.
Imagine the embarrassment when it's found out that the DNA in these bugs ISN'T made with As instead of P.
Dave is Wolfe-Simon? Looks like.
ReplyDeleteDave, Dr. Redfield just owned Wolfe-Simon, and everyone in the know agrees the Science paper was poorly done. Deal with it.
ReplyDeleteThere are plenty of scientists not involved with the work who have watched with dismay the way our scientific process and discussion has devolved. As a society we have become too accepting of the idea that every debate must be a cable news style shout fest where one attacks not only the messenger but everyone and everything associated with him/her, along with the message.
ReplyDeleteI'm not F.W.S., and for the record I agree with many of the criticisms leveled at the paper. However, I also understand that this is a preliminary report of what would be a very new phenomenon. The authors can't be expected to have searched down every possible avenue. They probably thought that they had enough to support and put forth their interpretation, and it would seem that the reviewers agreed. People could (and possibly will) spend years on further work. Really, the interesting question for everyone to take away from this is when do you have enough evidence to publish and when do you continue to hold off for more and different analyses? We all know that there is a rush to publish, and the authors may be guilty of "least publishable unit" syndrome here. Presenting enough to support their conclusion to reviewers and deferring other lines of investigation for future pubs. That was a big mistake for something of this magnitude. Its easy enough to make these points without turning in to Bill O'Riley and calling people pinheads.
Dear Dave, please try to remain objective and reasonable about this discussion. In this case, remaining objective means admitting that this study is a pile of unsubstantiated and contradictory ramblings.
ReplyDeleteA very strong response by other scientists was the only suitable course of action in this case, as it ensured that at least some of the criticism was picked up by the mainstream media. Claiming an illusory moral high ground from which to watch and be dismayed is naive at best; while a calm and measured discourse may be desirable and laudable, this is not how the world actually works, and will not get your voice publicly heard. This discussion is not about the scientific process or scientific publishing, but about the responsibilty of scientists to have the courage to stand up publicly to defend scientific integrity. I have no evidence to show that there are no intelligent woodlice on Mars, but that doesn't make it alright to go on TV and claim that I've found them.
That is pretty much what the authors did, or can you give an example of convincing evidence for P/As exchange? Just one?
-jj
After my early reaction (Arsenic and Odd Lace), I wrote a more extended critique of the successive gatekeeping failures that accompanied this paper, from the researchers to the reviewers to Science to NASA: The Agency that Cried "Awesome!"
ReplyDeleteAfter I had written it, the authors' FAQ appeared. Most of the items you list in this post jumped at me. Particularly egregious was the "PCR requires highly purified DNA" statement. It makes me wonder if they comprehend all the techniques they used as well as they seem to (not) comprehend this one.
To those who insist that Rosie is doing something inappropriate, think again. To let this paper sail by unchallenged would be a grave disservice to the scientific process. Let's just hope that the struggling discipline of astrobiology survives it.
Dave:
ReplyDelete"Shoutfests" are nothing new in science, occurring way before social media was invented. Of course they used to be limited by the physical constrains of the lab, conference room or hall.
Besides nucleic acids, there are other needs for P in a cell, in particular, for the phospholipids in a cell membrane. However, I believe that arsenolipids have been previously described in biology (in fish, such as cod), so if they were present in this bacterium, this could free-up some scarce P for nucleic acids. Also, as a microbiologist with some experience with electron microscopy, the "vacuoles" shown in the electron micrograph look to me more like granules of poly-beta-hydroxybutyrate (PHB), a carbon storage polymer. PHB is often formed when carbon is in excess and especially so when carbon is in excess and cells are growth-limited for some other nutrient. Perhaps P starvation has triggered PHB formation?
ReplyDeleteFWS is pushing herself out front on this.
ReplyDeleteI know a lot has been said about this, but I noticed something that may make the arguments about washing the DNA moot. Please bear with me; I predict that it will be worth it in the end.
ReplyDeleteThe critical data for the claim of As-DNA is in Table S2 (the numbers in Fig. 2A come from there). This table, as many have noted, raises all sorts of red flags. Why does the As-grown DNA band have less than half the P of the blank? Why does the P-grown RNA have three times less As than the blank? Given these facts, why should we make anything of the less than two-fold excess of As in the supposed As-DNA compared to the blank?
The error estimates in S2 seem to say that these differences are all real. But then why the large variation between things that should be the same? Where do these error estimates come from? I think I see where they come from for the blank, and the answer is disconcerting.
The P and As numbers for the blank on the left side of Table S2 appear to come from the results reported at the bottom of the "Blanks" section of Table S1. The units given are different, but the numbers make sense. For P, we see values of 795, 1066, and 599 for three replicates. The average of these numbers is 820, which appears in S2.
Where does the +/-143 in S2 come from? I think they got this by adding the errors in S1 (80, 107, 60) and dividing by sqrt(3). This is an odd computation in several ways. Critically, it takes no account of how different the measured values were. Given that S2 was supposed to contain twice the standard error, 143 is too low by almost a factor of two.
Now here's where it gets interesting. What about the 15 +/-3 value for As in the blank (S2)? In S1 they report values of 42.5, <2, and <2 for As. It seems that they took something like (42.5+2+2)/3 to get the 15. And then they came up with +/-3 by...I don't know. Just divided the +/-9 (for 42.5) by three? In any case, this grossly underrepresents the uncertainty.
It doesn't take a sophisticated statistical analysis to see the point. If my interpretation is correct, one of their three replicate measurements of the blank had greater than 50% more As than the supposed As-DNA band. If so, the higher measured As is the As DNA band (27 vs. 15) is utterly meaningless.
Can this really be what they did?
I understand that this study makes claims that if substantiated would fundamentally change our understanding of life, which gives it unusual weight. I also understand that there are methodological factors at issue which can only be fairly evaluated by peers.
ReplyDeletePoor selection of peers for the review? Your rebuttals have received so much attention, and perhaps deservedly so, but I have to question the widespread insinuations of wrongdoing, if only because they are so vehement. I understand that this study makes claims that if substantiated would fundamentally change our understanding of life, which gives it unusual weight.
ReplyDeleteRegards,
Cliff Merchant MD
The error estimates in S2 seem to say that these differences are all real. But then why the large variation between things that should be the same? Where do these error estimates come from? I think I see where they come from for the blank, and the answer is disconcerting.
ReplyDelete